
Вероятная причина колебаний численности…
© 2018. Люшвин П.В., Карпинский М.Г. ФГБНУ «Всероссийский научно-исследовательский институт рыбного хозяйства и океанографии» (ФГБНУ «ВНИРО»), Москва
Вероятная причина колебаний численности популяций каспийских и беломорских тюленей
Гипоксию и сокращение кормовой базы неоднократно пытались связать с изменениями стока рек, ледового покрова, антропогенными загрязнениями и другими факторами среды, однако сколь либо устойчивых и масштабных связей не обнаружено. Одной из причин масштабных неоднократных заморов бентоса и рыб предполагается выброс флюидов, в первую очередь метана, при активизации сейсмики. При дегазации метана, даже когда его концентрации менее 1 мг/л, происходит массовая гибель бентосных организмов. Бактериальное окисление метана приводит к гипоксии, заморам. Через 5-7 лет после интенсивной дегазации метана в местах зимовки тюленей (Северный Каспий и Белое море), происходит снижение их численности и добычи, что объясняется сокращением кормовой базы в период щенки и кормления, поскольку дегазация сопровождается заморными явлениями и гибелью бентоса.
Введение
Гипоксию резкие снижения биомассы макробентоса и заморы рыб неоднократно пытались связать с изменениями стока рек, ледового покрова и антропогенным загрязнением, однако связи не были однозначными, масштабными и продолжительными. Сопоставление перечисленных негативных явлений для организмов с активизацией региональных землетрясений даёт основание полагать о наличии взаимосвязей между ними. Формальным основанием для этого является то, что большинство гидробионтов в силу редкости землетрясений не адаптировано к литосферным флюидам, и присутствие метана даже в концентрациях менее 1 мг/л вызывает гибель молоди рыб [1]. Нахождение гидробионтов в районах разломов земной коры приводит к функциональным расстройствам и даже их гибели. Например, в кандалакшском разломе Белого моря биомасса и численность двустворчатых моллюсков в 2÷3 раза ниже, чем за пределами [2].
Снижение биомассы зообентоса в традиционных районах нагула сказывается на бентосоядных рыбах, поскольку их упитанность зависит от состояния кормовой базы, при уменьшении которой рыбы вынуждены искать пищу в других местах. Для сопоставления изменений упитанности рыб-бентофагов и сейсмострессов были выбраны нерестующие осетровые. У них отсутствует плавательный пузырь, поэтому они устойчивы к сейсмоакустическим воздействиям, а проводя часть жизни в пресных водах, с повышенным во время весенних паводков содержанием природных газов, резистентны и к метану [1]. Тюлени также устойчивы к газовому составу воды, поскольку дышат атмосферным воздухом. Однако, через 5-7 лет после землетрясений, произошедших в местах зимовок тюленей, снижается их численность и добыча, что можно объяснить резким уменьшением кормовой базы в период щенки и кормления.
Северный Каспий
В осадках Северного и Среднего Каспия местами наблюдаются аномально высокие содержания метана (1÷11 мг/кг), вследствие чего понижается биомасса бентосных организмов. Локальные максимумы площадей дефицита кислород, приуроченные к терскому берегу, в 1964, 1966, 1969, 1973 и 1981 гг. были связаны с весенне-летними землетрясениями (рис. 1) [3].
В годы резких снижений биомассы бентоса пищевые отношения между рыбами-бентофагами обостряются, конкуренция за пищу увеличивается, что сразу же влечет за собой снижение темпа роста, более высокое воздействие выедания на бентос. Так, например, в середине 50-х годов едва ли не весь кормовой бентос потреблялся рыбами-бентофагами (рис. 1, в). В годы сейсмострессовых воздействий на бентос (1954, 1956 гг.) у осетров и воблы в желудках было до 15-20 % растений и грунта, тогда как в сейсмоспокойные годы в 2-3 раза меньше. Лишь в 1957-1958 гг. нагрузка снизилась, причем произошло это на фоне ослабления сейсмострессовых нагрузок на бентос, а не за счет сокращения бентофагов. При сопоставлении изменений биомассы бентоса (с учетом его выедания бентофагами) с числом землетрясений в регионе оказалось, что рост биомассы совпал со снижением сейсмической активности в регионе [3].
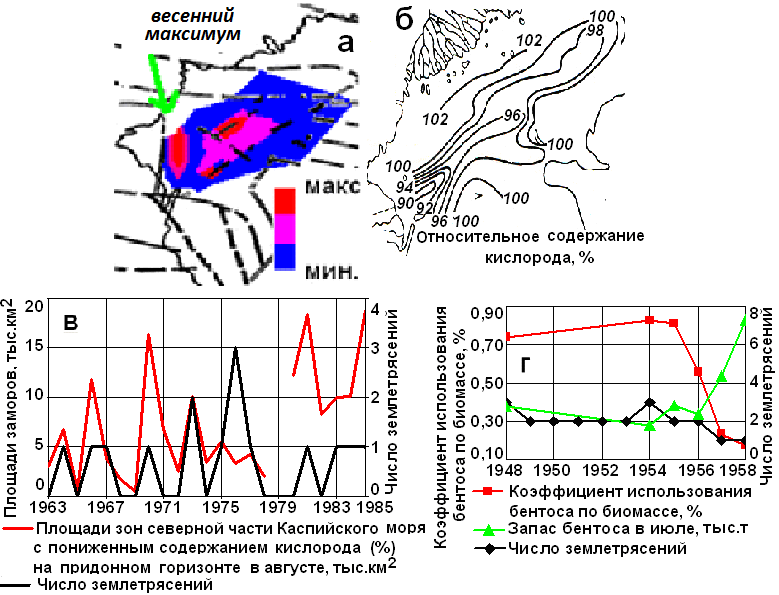
Рис. 1. Содержание метана в грунтах волжского взморья (а); среднее многолетнее (1961-1983 гг.) распределение относительного содержания кислорода в июне-июле, % (б); сопоставление биомассы митилястера в западной части моря, числа терских землетрясений и площади моря с пониженным содержанием кислорода в августе (в); коэффициент использования бентоса в Северном Каспии, запас бентоса в июле, число землетрясений (г).
«В 1988 г. отмечено катастрофическое снижение биомассы донной фауны в глубоководной части Северного Каспия. На этих обширных площадях осталось 1,5-2 % от общего количества донных животных в предшествующие и последующие годы. Аналогичное снижение было в 1938 г. при падении уровня моря. Резкое снижение количества бентоса нельзя объяснить интенсивным выеданием его рыбами, так как в этот период глубоководный район не являлся нагульным пастбищем моллюскоедов (осетра). Вероятно, это произошло из-за стабильных заморных явлений. По данным лаборатории водных проблем КаспНИРХа, насыщение воды кислородом у дна понижалось до 9 %. Площадь гипоксии в июне оказалась в 1,5 раза выше, чем за этот же период предыдущего года. В 1989 г. гидрологическая обстановка улучшилась, площадь гипоксии сократилась до 4,5 км2 и в бентосе пошёл в рост биомассы во всех группах беспозвоночных почти до уровня прошлых лет» [4, стр. 104]. Основные скопления осетровых в 1988 г. были в Южном Каспии. В 1989 г. ареал нагула осетровых увеличился до 28,0 тыс. км2, по сравнению с 19,3 тыс. км2 в 1988 г, количество осетровых в 1989 г. в Северном Каспии выросло почти вдвое [4].
Резкое снижение бентоса и необычное поведение осетровых в 1988 г. совпадает с активизацией землетрясений в зонах дагестанских и северокаспийских разломов (рис. 2). Единого отклика в развитии бентоса на эти землетрясения в солоноватых и пресных водах нет (рис. 2, а) [4]. Обусловлено это как мористым расположением активных разломов, так и влиянием на мелководный бентос ледовых явлений, речного стока, наносов, сгонов и нагонов. Лишь на глубинах свыше 10 м, где солёность воды превышает 4-6 ‰, усиление сейсмической деятельности сопровождается резким снижением биомассы бентоса – моллюсков, ракообразных, червей (рис. 2, б).
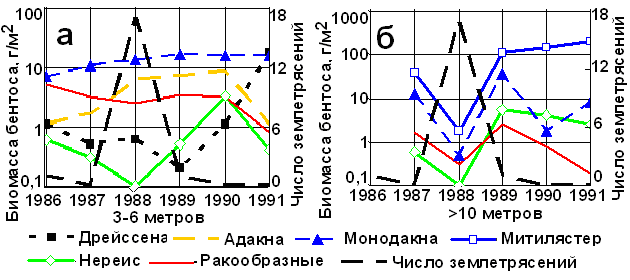
Рис. 2. Сопоставления числа терских землетрясений с биомассой моллюсков, нереиса и ракообразных на глубинах 3÷6 м (а) и свыше 10 м (б).
Жизнедеятельность каспийских тюленей (Pusa caspica) во многом зависит от сейсмоактивности (рис. 3). Через 5-7 лет после роста числа землетрясений падает добыча тюленей. Объяснить это можно влиянием снижения кормовой базы – крабов, мизид и бычков, активно потребляемых тюленями в зимний период, когда происходит щенка и выкармливание [3]. В результате через 5-7 лет появляется малочисленное поколение. Так и случилось через 5-6 лет после обсуждаемого выше сейсмострессового для бентоса 1988 г. В 1994 г. добыча тюленя упала более чем вдвое по сравнению с 1988-1993 гг.
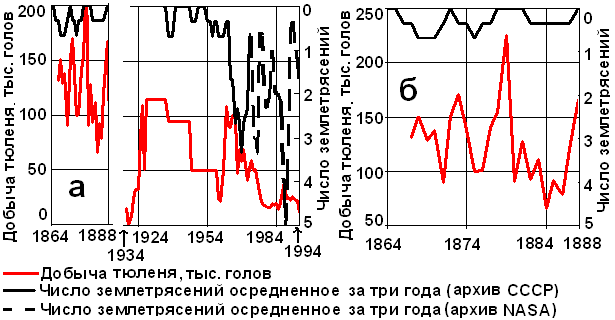
Рис. 3. Сопоставление добычи тюленя в Каспийском море и сдвинутого на 5 лет осреднённого за три года числа землетрясений в северо-каспийском регионе (а); увеличенный фрагмент рис. А (б).
Белое море
Уловы (воспроизводство) наваги (Eleginus nawaga) в Белом море лимитируются сейсмической активностью (рис. 4). Через год-два после учащения землетрясений в её уловах наступают локальные минимумы. В сейсмоспокойные годы от 70 до 100 % рыб готовы к нересту в двухгодовалом возрасте. В сейсмоактивные годы их число снижается до 30 %. Резкие спады численности рыб – кормовой базы гренландских тюленей (Pagophilus groenlaudica) в период щенки, кормления и питания молоди, также как и в Каспии через 6-8 лет приводят к обвалам боя тюленей [5].
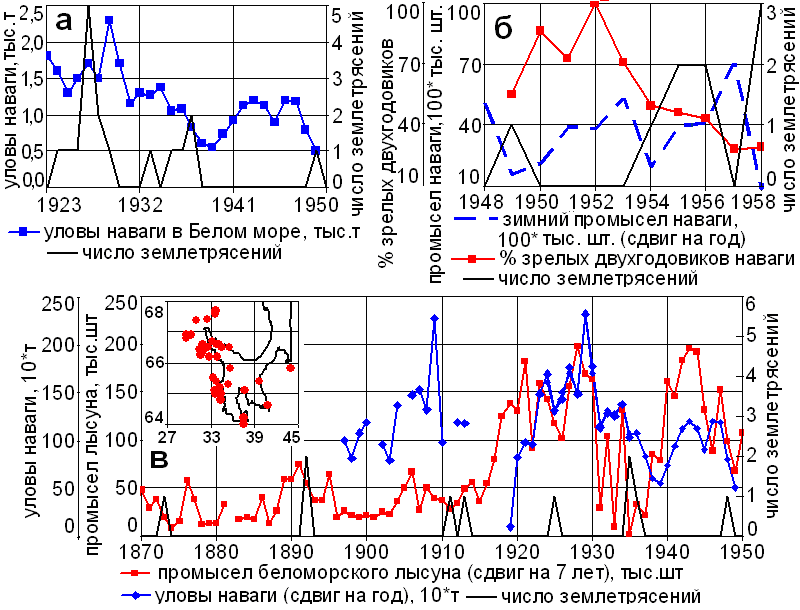
Рис. 4. Сопоставление числа землетрясений с уловами наваги, % зрелых двухгодовиков наваги и промыслов беломорского тюленя (на врезке эпицентры землетрясений в Белом море).
Выводы
Приведённый материал показывает, что при активизации землетрясений угнетается бентос, снижается упитанность рыб-бентофагов и посещение ими этих акваторий, даже если они являются традиционно нагульными. Когда такие процессы происходят в местах зимовки тюленей, то из-за резкого снижения кормовой базы в период щенки и кормления через 5-8 лет резко снижается численность и добыча тюленей. Наблюдается тенденция развития гипоксии в придонных слоях воды при активизации землетрясений.
Литература
1. Патин С.А. Экологические проблемы освоения нефтегазовых ресурсов морского шельфа. М.: Изд. ВНИРО, 1997. 349 с.
2. Ромадоновская М.А., Зайцев В.А., Малютин О.И. Новейшие разломы как трендовый геопатогенный фактор воздействия на развитие бентосных сообществ Кандалакшского залива Белого моря // Система планета Земля. Нетрадиционные вопросы геологии. Материалы научных семинаров. М. 1999. С. 218-224.
3. Люшвин П.В., Карпинский М.Г. Причины резких сокращений биомасс зообентоса и их последствия // Рыбное хозяйство. 2009. №5. С.65-69.
4. Смирнова Л.В. Состояние зообентоса Северного Каспия в конце 80-х годов // Морские гидробиологические исследования. М.:ВНИРО. 2000. С. 103-110.
5. Люшвин П.В. Заклепки на Науках о Земле. Результат возведения парадигм в статус аксиом в геофизике и гидробиологии // М. издание автора, 2016. 147с. Стационарный сетевой адрес: http://nadisa.org/download/lyushvin_book.pdf.